Вегетативната нервна система регулира функции, които са характерни и за растителния свят - обмяна на веществата и размножаване. Тя инервира вътрешните органи, жлезите, сърцето и кръвоносните съдове. Тази инервация е моторна и сетивна. Функциите на вегетативната нервна система са рефлексни и не се регулират от волята на индивида. Поради това много често тази нервна система се нарича и автономна.
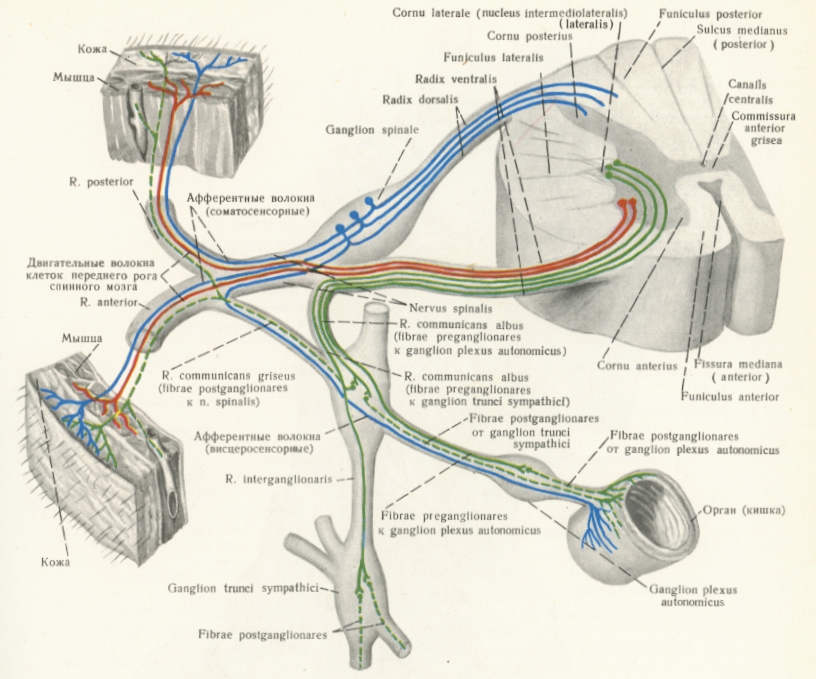
Вегетативната нервна система се състои от централна и периферна част. Централната част включва вегетативни ядра, които са разположени в гръбначния мозък и мозъчния ствол. Хипоталамусът регулира важни вегетативни функции на по-високо ниво, поради което Шерингтон го нарича главно ядро на вегетативната нервна система. Периферната част на този дял на нервната система се състои от възли, нерви и сплетения. Основна разлика между вегетативната и соматичната нервна система е в устройството на еферентната част на рефлексната дъга. При соматичната нервна система еферентното рамо е едноневронно и включва мотоневроните в предните колонки на сивото вещество на гръбначния мозък и в моторните ядра на черепномозъчните нерви в мозъчния ствол. При вегетативната нервна система еферентното рамо е двуневронно. Перикарионът на първия еферентен неврон е разположен във вегетативни ядра в латералния стълб в сивото вещество на гръбначния мозък и във вегетативните ядра на черепномозъчните нерви в мозъчния ствол. Аксонът на неврона достига до периферен вегетативен възел. Поради това първият неврон се нарича предвъзлов. Във вегетативните възли се намират телата на нервни клетки, които са вторият еферентен неврон. Аксоните им достигат до ефекторните органи. Този втори неврон се нарича следвъзлов. Във възлите се образуват синапси между пред- и следвъзловите неврони, като една предвъзлова контактува с много следвъзлови нервни клетки - дивергентен тип на предаване на информацията. Аферентното рамо на вегетативната нервна дъга съвпада напълно с това на соматичната нервна система. Аферентните неврони са малките псевдоуниполарни клетки в спиналните възли или сетивните възли на черепномозъчните сетивни ядра.
Схема на рефлексната дъга на симпатикусовата нервна система. Със зелено са означени симпатикусовите нервни влакна, със синьо - сетивните влакна, а с червено - двигателните. Двигателните нерви не се прекъсват в междинни ганглии - от ядрата в гръбначния мозък те достигат директно до ефекторните органи - най-вече мускули. Еволюционно живите организми са изградени по такъв начин, за да има бързина в действията (най-вече при тези, които зависят от волята на организма) - което е необходимо в много случаи за оцеляването на индивида. Същевременно много вегетативни функции се регулират от периферните ганглии на симпатикусовия и парасимпатикусовия дял - по този начин се освобождават по-висшите дялове на нервната система и те използват своя капацитет за по-висши функции от по-голямо значение. Делегирането на права и задължения (принцип в съвременния мениджмънт) е измислено от Природата много преди хората да започнат да основават и развиват компании! Организмът на бозайниците е една многопластова структура с много нива на управление и контрол - съществуват мениджъри от ниско, средно, високо и най-високо ниво, докато по-нисшите организми са по-плоски управленски структури - при тях нивата на мениджмънт са доста по-малко и не се налага делегиране на функции на по-нисшестоящите в йерархията.
Съществуват и други разлики между вегетативната и соматичната нервна система. Вегетативните нерви са с тънка миелинова обвивка или са амиелинови, поради което провеждат нервния импулс с по-малка скорост. Вегетативната нервна система осъществява двустранна инервация на вътрешните органи, поради което е дифузна и недобре локализирана. Вегетативните пътища в централната нервна система са недобре обособени морфологично.
Вегетативната нервна система се разделя на два дяла - симпатикова и парасимпатикова. Критериите за това разделяне са морфологични, функционални и нервнотрансмитерни. Според морфологичните критерии при симпатикусовия дял предвъзловите еферентни неврони са разположени в substantia intermediolateralis на латералното рогче на гръбначномозъчните сегменти от C8 до L2. Поради това този сегмент се нарича тораколумбална част на ВНС. При парасимпаткуса предвъзловите еферетни неврони се намират в ядрата на черепномозъчните нерви в мозъчния ствол, както и в латералния рог на сегменти S2 - S4 на гръбначния мозък, поради което тези части се наричат краниосакрална част на ВНС. Периферните вегетативни възли на симпатикусовия дял са разположени около гръбначния стълб - паравертебрални и превертебрални, близо до гръбначния мозък. При парасимпатикуса възлите се намират близо до органите, които инервират, а понякога и вътре в стената им, поради което въпросните възли се наричат терминални пара- или интраорганни. При симпатикусовия дял има къси предвъзлови и дълги следвъзлови нервни влакна, докато при парасимпатикусовия дял е точно обратното - следвъзловите са много къси, понякога микроскопични. Този критерий е функция от разположението на ганглиите. Освен това при симпатикуса има силно изразена дивергенция - средното съотношение е един предвъзлов неврон да се свързва с около 200 следвъзлови. При парасимпатикусовия дял също има дивергенция, но съотношението е само 1:20.
Названието симпатикусов дял е дадено от древни изследователи - още преди няколко хиляди години на хората е направило впечатление че симпатикусовите нервни влакна буквално симпатизират на други нерви - най-често двигателни. Ходът на тънкото амиелиново влакно е винаги успореден на дебелото миелиново влакно на двигателния нерв. В даден момент обаче се налага разделяне - двигателният нерв се насочва към мускул, а вегетативният - към жлеза или вътрешен орган. Тогава симпатикусовият нерв намира друга структура, към която да се присъедини - кръвоносните съдове. Около тях се образуват сплетения, които достигат до всеки един орган с необходимост от вегетативна инервация.
Съществува и функционален критерий за различаване на двата дяла на вегетативната нервна система. При симпатикуса реакциите са по-дифузни, докато при парасимпатикусовия дял са по-локализирани. Това се обуславя от дивергенцията при контактуването на предвъзловите със следвъзловите неврони. Освен това двете части оказват противоположен ефект при инервираните органи. Действията на симпатикуса са свързани с изразходване на повече биологична енергия - този дял се включва при необходимост от преодоляване на силни или вредни въздействия върху организма - бягай или се бий! Обменните процеси са ускорени, като преобладават катаболитните. Действията на парасимпатикуса са свързани с преобладаване на анаболитните процеси - прием на хранителни вещества и съхраняване на енергия.
Преганглионерните неврони на симпатикуса са холинергични, а постганглионерните - адренергични. Изключение правят постганглионерните влакна на потните жлези, които са холинергични. При парасимпатикусовата вегетативна нервна система преганглионерните и постганглионерните неврони са холинергични.
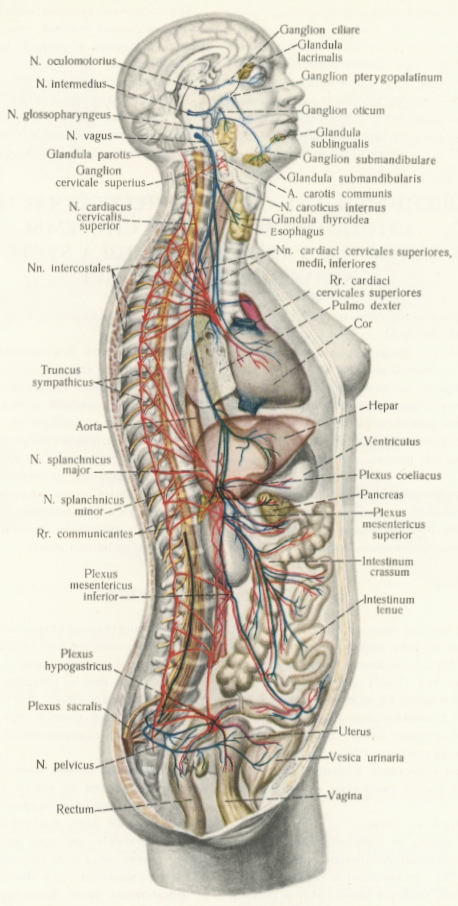
Обща схема на всички вегетативни нерви в организма. Симпатикусовите влакна са отбелязани с червено, а парасимпатикусовите - със синьо. Вижда се значително по-големият обем на симпатикусовата нервна система.
Вегетативната инервация на лицево – челюстната област се осигурява от различни черепномозъчни нерви. Основна роля играят n. facialis и n. glossopharyngeus; освен тях парасимпатикусови влакна влизат в състава на n. occulomotorius и n. vagus. Те пренасят парасимпатикови нервни влакна, които се прехвърлят към други нерви (главно сетивни) и така достигат до различни структури, които инервират. Симпатиковата инервация на областта се осигурява от горния шиен симпатиков ганглий, от който по хода на различни кръвоносни съдове достигат симпатикови нерви.
Парасимпатикусовото ядро на n. occulomotorius е nucleus occulomotorius accessorius (ядро на Edinger - Westphal), което е разположено в tegmentum mesencephali. Аксоните на нервните му клетки съставляват парасимпатикусовата част на нерва, преминават в неговия долен клон и достигат до ganglion cilliare, разположен в очницата. Следвъзловите влакна на нерва формират късите цилиарни нерви, които достигат до очната ябълка и инервират цилиарния мускул и сфинктера на зеницата - поради което и при повишен парасимпатикусов тонус са налице свити зеници.
Парасимпатиковото мозъчно ядро на лицевия нерв се състои от разсеяни неврони в дорзолатералната част на мостовата ретикуларна формация. Неговите граници се преливат в заобикалящата го тъкан и са лошо очертани. Всъщност преганглионерните парасимпатикови неврони на това ядро представляват ростралната част от непрекъсната верига, която се простира от продълговатия мозък до моста. В нейния каудален край са разположени клетките, образуващи дорзалното ядро на блуждаещия нерв. В моста клетките на тази колона достигат точно до връзката продълговат мозък – мост и каудалния полюс на моторното ядро на лицевия нерв. Преганглионерните парасимпатикови влакна (аксони) от nucl. salivatorius rostralis образуват в периферията n. intermedius, който скоро след излизане от мозъчния ствол се присъединява към лицевия нерв. После парасимпатикусовите влакна напускат ствола на n. facialis в състава на n. petrosus major и chorda tympagni. N. pеtrosus major е първият клон на лицевия нерв и преминава през hiatus et sulcus n. petrosi majoris към foramen lacerum, пробива fibrocаrtilago basilaris, присъединява се към симпатиковия n. petrosus profundus и заедно образуват n. canalis pterygoidei. Вкусовите влакна в състава на нерва преминават без прекъсване в парасимпатиковия ганглий и достигат небцовите нерви. N. petrosus profundus е образуван от симпатиковите влакна около a. carotis interna. Двата нерва достигат до ganglion pterygopalatinum. Последният е най-големият от периферните парасимпатикови ганглии. Разполага се в дълбочината на fossa pterygopalatina близо до foramen sphenopalatinum и пред canalis pterygoideus. Той лежи точно под n. maxillaris, където последният пресича криловиднонебцовата яма. Горночелюстният нерв отделя две малки сетивни клончета към ганглия, които осигуряват сетивната инервация на регионите, инервирани от клоновете на ганглия. Парасимпатиковите влакна на n. petrosus major образуват синаптични контакти с клетките на ганглия. Постганглионерните парасимпатикови влакна преминават към ябълчния нерв и оттам чрез анастомоза към слъзния нерв осигуряват инервацията на секреторно – моторната дейност на слъзната жлеза. Останалата част от влакната влизат в състава на небцови, носни и гълтачни клончета, които регулират дейността на съответните малки жлези. От своя страна постганглионерните симпатикови клончета в състава на n. petrosus profundus регулират вазомоторната дейност на региони, снабдявани от съответните нервни клонове. Като обособени клонове от дисталния край на ganglion pterygopalatinum, в които участват симпатикови и парасимпатикови нервни влакна, са следните нерви:
Rami orbitales са 2 – 3 клончета, които навлизат в орбитата през fissura orbitalis inferior и снабдяват периоста, а през foramen ethmoidale posterior снабдяват клиновидния и решетъчния синус
Nn. palatini са три клона. N. palatinus major навлиза в canalis palatinus major и излиза през големия небцов отвор, след което в бразди на твърдото небце (3 – 4 клончета) стига до гингивата на премоларите и моларите от палатиналната им страна, лигавицата и жлезите на твърдото небце (последните имат най-вече парасимпатикусова инервация). B canalis palatinus major нервът отделя няколко клончета, които през перпендикулярната плочка на небцовата кост проникват и инервират долната носна мида и стените на средния и долния носен ход. Nn. palatini minores, напускайки небцовия канал, преминават през малки небцови отвори и инервират мъжеца, небцовите сливици и мекото небце
Rami nasales навлизат в носната кухина през foramen sphenopalatinum, образувайки латерална и медиална група. Латералната група инервира задните части от лигавицата на горната и средната носна мида, както и на задните решетъчни кухинки. Медиалната група нервни клончета инервират лигавицата от задните части на носния покрив и носната преграда. Сред тези нерви се откроява по-дълъг и дебел клон, n. nasopalatinus, който преминава в предно – долно направление по носната преграда в улей на ралника и през canalis incisivus достига предната част на твърдото небце. Той инервира предните части на носната преграда и анастомозирайки с предните небцови нерви, участва в инервацията на лигавицата от предната част на твърдото небце
Ramus pharyngeus преминава през canalis palatovaginalis заедно с фарингеалния клон на а. maxillaris и инервира лигавицата на назофаринкса зад отвора на слуховата тръба.
Chorda tympani напуска лицевия нерв на около 6 mm. преди foramen stylomastoideum и пробива задната стена на тъпанчевата кухина близо до задния ръб на тъпанчевата мембрана. Минавайки между фиброзния и лигавичния слой на мембраната, тя излиза през fissura petrotympanica и се спуска вентрално към медиалната повърхност на spina ossis sphenoidalis и се насочва към m. pterygoideus lateralis. Нервът се прибавя под остър ъгъл към n. lingualis. Чрез този нерв се доставят преганглионерни парасимпатикови влакна на ganglion submandibulare, както и аферентни влакна, носещи импулси от вкусовите луковици по предните 2/3 на езика. Парасимпатикусовите влакна достигат подчелюстния ганглий и осъществяват синаптичен контакт с клетките му.
Ganglion submandibulare е малък вретеновиден парасимпатиков ганглий. През ганглия преминават и постганглионерни симпатикови влакна от сплетението около a. facialis, които вървят като израстъци на клетки в горния шиен симпатиков възел. Постганглионерните парасимпатикови влакна заедно с постганглионерните симпатикови влакна влизат в състава на n. lingualis. Чрез клончетата на езичния нерв към подчелюстната, подезичната и предните малки езикови жлези се осигурява тяхната секреторно - моторна регулация и вазомоторната регулация на тъканите в областта.
Инервация на лицево - челюстната област Лицево - челюстна хирургия Зъбни импланти
N. glossopharyngeus също осигурява част от парасимпатиковата инервация на лицево – челюстната област. Той води началото си от две ядра в продълговатия мозък (моторно и парасимпатиково). Nucleus salivatorius caudalis е висцерално моторно ядро на езиковогълтачния нерв. Съдържа мултиполярни нервни клетки. Разположено е в дълбочината на продълговатия мозък като продължение на горното саливаторно ядро и в същото време е близо до дорзалното ядро на блуждаещия нерв. Според някои автори това ядро се приема като челна група, еквивалентна на nucleus dorsalis nervi vagi. Да се определи и отграничи клетъчната му популация от тази на ретикуларната формация е изключително трудно. От него излизат преганглионерни парасимпатикови моторни влакна, минават през n. petrosus minor и достигат ganglion oticum по хода на n. tympanicus. Последният се отделя от ganglion inferius n. hypoglossi и навлиза в тъпанчевата кухина през canaliculus tympanicus, като образува plexus tympanicus. Освен аферентни влакна сплитът съдържа симпатикови нервни влакна чрез анастомозата с горния шиен симпатиков възел, достигащи до тук по хода на средната менингеална артерия, а също и парасимпатикови преганглионерни нервни влакна. Двата вида вегетативни влакна се обособяват самостоятелно като n. petrosus minor. Последният преминава от каналче под полуканала на m. tensor tympani и достига предната повърхност на пирамидата, където от собственото си каналче преминава през synchondrosis sphenopetrosa, за да достигне ganglion oticum.
Ganglion oticum е малък, овално – плосък, сиво – червеникав периферен парасимпатиков ганглий, разположен топографски под foramen ovale от медиалната страна на n. mandibularis. Симпатиковите постганглионерни влакна преминават през ганглия, без да образуват синаптични контакти с клетките му. Двата вида влакна осъществяват секреторно – моторната и вазомоторната регулация на задушната жлеза. Те се прибавят към n. auriculotemporalis още при изграждането му от две клончета, които заграждат а. meningea media. След това нервът преминава между ligamentum sphenomandibulare и шийката на долната челюст и се насочва латерално зад долночелюстната става към горната част на задушната жлеза (където се отделя клонче за нейната парасимпатикова инервация) и към повърхностните слепоочни съдове. Тук n. auriculotemporalis пресича ябълчния израстък на слепоочната кост и изпраща последните си разклонения към кожата и фасцията на слепоочието и скалпа.
Парасимпатикусов компонент има и блуждаещият нерв. Неговото ядро е nucleus dorsalis nervi vagi, което е разположено в продълговатия мозък. Предвъзловите влакна достигат до ганглии, разположени в непосредствена близост или в органите, инервирани от нерва - терминални ганглии. Следвъзловите влакна са къси и инервират органите в областта на шията, гръдната кухина и част от тези в коремната кухина - бъбреци, пикочопроводи, задстомашна жлеза, стомах, тънко черво и дебело черво до flexura colli sinistra.
Симпатикусовата част е по-големият дял от вегетативната нервна система. Тя е съставена от два ганглийни ствола, симпатикусови нерви, сплетения и възли. Предвъзловите неврони на симпатикуса се намират в substantia intermediolateralis на гръбначномозъчните сегменти от С8 до L2. Поради това се нарича тораколумбален отдел на ВНС. Миелиновите следвъзлови аксони напускат гръбначня мозък в състава на предните коренчета на спиналните нерви и като rami communicantes albi достигат възлите на симпатикусовия ствол. Възможно е те да преминат транзитно през тях и да достигнат до превертебралните възли в симпатикусовите сплетения. Аксоните на следвъзловите неврони са амиелинови и отиват в две направления - като rami communicantes grisei се присъединяват отново към спиналните нерви и достигат до ефекторните органи или да сформират самостоятелни нерви, които се оплитат около кръвоносните съдове и така достигат до ефекторните органи.
Симпатикусовият ствол е чифтен, разположен е от двете страни по протежение на гръбначния стълб. Представлява верига от 22 - 23 възела, които са свързани помежду си с междувъзлови нервни влакна. Различават се шийна част с 3 възела, гръдна част с 10 - 12 възела, поясна (коремна) част с 4 възела и тазова част с 4 възела. Ганглиите на симпатикусовия ствол получават предвъзлови влакна от rami communicantes albi. Последните достигат само до гръдните и първите два поясни възела. Шийните възли получават предвъзлови влакна от осми шиен и първите три гръдни сегмента на гръбначния мозък. Като бели комуникационни влакна те влизат в съответните ганглии на гръдната част, но не прекъсват в тях, а продължават възходящо като rami interganglionares и достигат и прекъсват в трите шийни лимфни възела. Трети и четвърти поясен и всички тазови възли получават предвъзлови влакна от долните гръдни и първите два поясни гръбначномозъчни сегмента. Тези влакна в състава на rami communicantes albi влизат в съответните възли на симпатикусовия ствол без да прекъсват в тях, спускат се надолу по хода на интерганглионерните влакна и достигат до поясните и тазовите възли. Следвъзловите влакна излизат от симпатикусовия ствол като rami communicantes grisei към спиналните нерви или самостоятелно към ефекторните органи. В тях се съдържат и висцеросетивни влакна, които идват от спиналните нерви, преминават през rami communicantes albi и транзитно през възлите на ствола.
Шийната част на truncus sympaticus се състои от три възела - горен, среден и долен. Разполагат се пред напречните израстъци на шийните прешлени, под lamina prevertebralis на шийната фасция, зад вътрешната сънна, респективно общата сънна артерия. Горният шиен възел се намира на нивото на втори - трети шиен прешлен, има вретеновидна форма и дължина около 3 - 4 см. От него излизат следните нерви:
n. jugularis - той се изкачва до основата на черепа и се разделя на две клончета - едното е свързано с долния ганглий на езиковогълтачния нерв, а другото - с горния ганглий на блуждаещия нерв. Този нерв отделя клончета към bulbus jugularis superior, където те се свързват с glomus caroticus
n. caroticus internus върви заедно с вътрешната сънна артерия, разпада се на клончета и образува около артерията сплетение - plexus caroticus internus. Това сплетение е най-гъсто когато артерията навлиза в кавернозния синус, където се образува plexus cavernosus. От него излизат вторични сплетения около клоновете на вътрешната сънна артерия - plexus ophtalmicus, plexus arteriae cerebrae anterioris и plexus arteria cerebri mediae. Освен това се отделят свързващи клончета към n. occulomotorius, n. trochlearis и n. ophtalmicus. От последния нерв клончетата достигат през n. nasociliaris и nn. ciliares longi до m. dilatator pupillae. Предвъзловите влакна за този мускул произхождат от centrum ciliospinale, разположен в първия гръден сегмент на гръбначния мозък. Освен това plexus caroticus internus отделя nn. caroticotympanici в каротидния канал, които през едноименни каналчета проникват в средното ухо и се присъединяват към plexus tympanicus, участвайки в инервацията на лигавицата на тъпанчевата кухина и слуховата тръба. N. petrosus profundus достига до птеригопалатиновия ганглий като негов симпатиков корен, преминава без прекъсване през възела и достига слъзната жлеза и жлезите на небцето и носната кухина.
nn. carotici externi са два или три клона, които образуват сплетение около външната сънна артерия. Това сплетение продължава по хода на клоновете на тази артерия и отделя симпатикови корени към ganglion oticum за инервация на паротиднаа жлеза и ganglion submandibulare за инервация на субмандибуларната и сублигвалната жлеза. Симпатикусовите корени преминават през възлите без да се прекъсват в тях.
rami laryngopharyngei достигат до латералната повърхност на гълтача и участват в образуването на вегетативното сплетение plexus pharyngeus. От него се отделят клончета за гръкляна и гълтача.
n. cardiacus cervicalis superior върви низходящо зад общата сънна артерия, достига до аортната дъга и под нея влиза в състава на plexus cardiacus
rami communicantes grisei са четири клончета към първите четири шийни спинални нерва
Средният шиен възел (ganglion cervicale medium) се разполага на мястото, където truncus thyrocervicalis кръстосва симпатикусовия ствол. Това е нивото на четвъртия шиен прешлен. Този възел е най-малък по размер и често е вариабилен. От него излизат nervus cardiacus cervicale medius, един клон към щитовидната жлеза и няколко малки комуникантни клончета към пети и шести шиен нерв. Кардиачният клон върви зад общата сънна артерия и зад подключичната артерия и под аортната дъга навлиза в plexus cardiacus.
Долният шиен симпатикусов възел се разполага под напречния израстък на седми шиен прешлен и в 75 % от случаите е срастнал с първия гръден възел - при което се образува ganglion cervicothoracicum (понякога наречен ganglion stellatum поради звездовидната му форма). Каудално ганглият достига до главата на първо ребро зад подключичната артерия. Възелът е обект на медикаментозни блокади при упорити вегетативни болки. От възела се отделят:
n. cardiacus cervicalis inferior - той преминава зад truncus brachiocephalicus вдясно или зад общата сънна артерия вляво. Низходящо достига до аортната дъга и зад нея навлиза в plexus cardiacus
rami communicantes grisei са два - за седми и осми спинален нерв
клонове, образуващи сплетения около a. subclavia a. vertebralis
Ansa subclavia е примка, която обхваща a. subclavia и се образува от rami interganglionares. Те вървят от долния към средния ганглий на симпатикусовия ствол. Това са предвъзлови влакна, произхождащи от centrum ciliospinale в първи гръден сегмент на гръбначния мозък, навлизащи в първи гръден възел без да се прекъсват в него. Насочват се възходящо по ствола като rami interganglionares и достигат първи шиен възел. Тук образуват синапси със следвъзловите неврони, които изпращат аксоните си за инервиране на гладките мускули - m. dilatator pupilae, m. tarsalis superior (Muller) и m. orbitalis. При увреждане на ansa subclavia тези мускули отпадат от функция и се развива синдромът на Клод Бернар и Хорнер - птоза (спускане на клепача), миоза (свита зеница) и енофталм (хлътване на очната ябълка).